Presentation Points:
- We have revised some of the basic frameworks considered for the control of the development of leaf primordia (Note 1) in plants and constructed an abstract model that can reproduce almost all of the regular patterns seen in leaf arrangement (the phyllotaxy (Note 2)).
- We have solved some of the remaining problems in the phyllotaxis, such as the emergence of the coccolithophorid-type phyllotaxis (Note 3), which has no symmetry, and clarified the importance of temporal variation in the interplastidial action in the control of leaf primordial development.
- It is expected to lead to the elucidation of the beautiful geometric patterns woven by nature from a generative principle.
Abstract:
The leaf primordium of plants arises at the periphery of the meristematic tissue (Note 4) at the stem apex and develops into a regular pattern of arrangement around the stem called the phyllotaxis. In most plants, the phyllotaxy is either spirally symmetrical (Note 5) or radially symmetrical, and can take one of the following patterns: double-row alternating (Note 6), Fibonacci spiral (Note 7) (a spiral whose opening (Note 8) is close to the golden angle (Note 9), crossed opposite (Note 10), or trilobed (Note 11). These major phyllotaxis patterns have been explained by the inhibitory field model, which assumes that new phyllotaxis arises to avoid the inhibitory effect of phyllotaxis formation exerted by the existing phyllotaxis. However, there are some phyllotaxis that cannot be explained by the conventional model. A typical example is the coccolithophoridian phyllotaxis, which has no symmetry and changes its opening periodically from 180° to 90° to 180° to 270°. A group led by Graduate Student Takaaki Yonekura and Associate Professor Munetaka Sugiyama of the Graduate School of Science at the University of Tokyo, Associate Professor Akitoshi Iwamoto of Kanagawa University (at the time of their research at Tokyo Gakugei University), and Assistant Professor Hironori Fujita of the Astrobiology Center and National Institute for Basic Biology extended the inhibitory field model by introducing a protoplast age-dependent variation in inhibitory force, This extended model also better matches the actual phyllotaxis than the conventional model in other respects. These results demonstrate the importance of temporal variation in interplastron action and contribute to an essential understanding of the phyllotaxis, which is expected to reveal the beautiful geometrical patterns of nature from a generative principle.
Presentation Description:
The style of arrangement that plant leaves draw around the stem is called the phyllotaxis. In most plants, the phyllotaxis exhibits either helical or radial symmetry that satisfies certain mathematical rules, and most of them follow one of the following patterns: double-row alternating, Fibonacci spiral (a spiral with opening degrees close to the golden angle), crossed opposite, or trilobate. The regularity of these patterns has long been a source of interest and research. Already in the 19th century, morphological observations pointed out that when a new leaf primordium arises at the periphery of the apical meristem, it seems to choose the position that maximizes its distance from the preceding primordium. This was the first time that a new leaf primordium had been observed. Subsequently, it came to be thought that the position of the new protoplast emergence is determined under the influence of inhibitory protoplast formation from the preceding protoplast, which shapes the pattern of the phyllotaxy, and a number of mathematical models were proposed that attempt to explain the phyllotaxy based on this idea. Among the most notable of these models are the two suppression field models by Douady and Couder: DC1, which specializes in alternate phyllotaxy, and DC2, which deals with the entire phyllotaxy.
Both DC1 and DC2 assume that each leaf primordium exerts a constant inhibitory force on primordium formation that decays with distance, and in addition to this, DC1 assumes that one new primordium is generated at the periphery of the meristem at each defined time point where the sum of the inhibitory effects from the preceding primordium is lowest, and Douady and Couder found that computer simulations using DC1 generated double-row reciprocal and Fibonacci helices depending on the parameters. Douady and Couder showed that computer simulations using DC1 produced double-row reciprocal and Fibonacci helices, while simulations using DC2 produced all four major phyllotaxis patterns, indicating that the suppression of new protoplast formation by the preceding protoplast is the determining factor in the phyllotaxis. Although this suppression was hypothetical in the DC model, recent physiological and molecular biological analyses have revealed that the plant hormone auxin is essential for the generation of new leaf primordia and that the preceding primordia absorb auxin from their surroundings, thus allowing us to capture the substance of the suppressive action as auxin deprivation. This has led to the realization that the substance of the inhibitory action can be captured as auxin deprivation. Following a series of such studies, the framework for controlling phloem primordial development assumed by the DC model is now widely accepted as the basic mechanism of phloem patterning.

Branch and winter bud of P. kokusagi. The winter buds were observed by scanning electron microscopy after removing the leaf primordium before P3 (O is the center of the apical meristem, P is the leaf primordium, and the smaller the number, the younger the shoot). The characteristic of the coccolithophorous phyllotaxy, i.e., alternation with periodic changes in openness, can be seen.
However, the DC model did not explain all the phyllotaxis patterns, and some remained unexplained. A typical example is the kokusagi-type phyllotaxy, which has neither spiral nor radial symmetry and changes its opening periodically from 180° to 90° to 180° to 270°. The kokusagi-type phyllotaxy is found in a small number of taxa, such as the kokusagi (Fig. 1) of the Mucoraceae and the shagmal lily (Liliaceae) of the Cyperaceae, suggesting that the kokusagi-type is not the result of a specialized framework for controlling leaf base development itself, but a common framework that produces the kokusagi-type under specific conditions. If this is the case, then the conventional model is likely to produce the kokusagi-type. If this is the case, then the inability of the conventional model to generate a kokusagi-type phyllotaxy indicates that the assumptions of the model are inadequate.
Based on the above idea, a group led by Graduate Student Takaaki Yonekura and Associate Professor Munetaka Sugiyama of the University of Tokyo Graduate School of Science, Associate Professor Akitoshi Iwamoto of the Kanagawa University Faculty of Science (at the time of the research, Faculty of Education, Tokyo Gakugei University), and Assistant Professor Hironori Fujita of the Astrobiology Center and Institute for Basic Biology, has reviewed the framework for controlling leaf primordial development They attempted to extend the DC model to generate almost all phyllotaxis patterns, including the coccolithophorid-type phyllotaxis. The focus was on the inhibitory effect of the leaf primordium on primordial formation. In conventional models, this is assumed to be of constant strength from the moment the primordium appears, which is a rather unnatural assumption from a biological point of view. Therefore, we introduced a time-dependent change in this inhibitory force depending on the age of the protoplast and constructed EDC1, an extension of DC1, and EDC2, an extension of DC2. Using these models, we ran exhaustive computer simulations and found that when the inhibitory force increases slowly in EDC2, a pattern of coccolithophoridian phyllotaxis is generated under certain parameter conditions (Fig. 2). This cockscomb-type phyllotaxy was found to be in close proximity to Fibonacci helices and cross pairs in parameter space (Note 12), and at the boundaries, the symmetry of the generated pattern appeared and disappeared with slight changes in parameters.
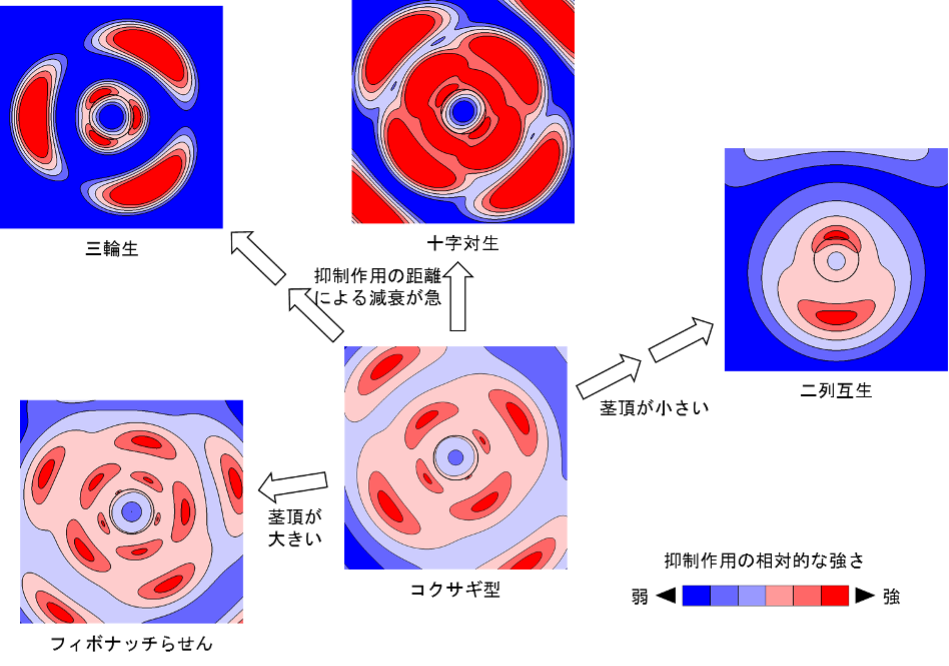
Under conditions of relatively slowly increasing inhibition, EDC2 computer simulations produced a cockscomb-type phyllotaxy in addition to the main phyllotaxy. Each panel shows the phyllotaxis pattern as a contour plot of the inhibitory effect, with the stem apex viewed from above.
EDC2 also found that the frequency distribution of the phyllotaxis better reflects reality than previous models. In real plants, the majority of helical phloem openings are close to the golden angle, but simulations of conventional models tend to produce helical phloem openings between 180° and the golden angle, which does not fully explain the overwhelming dominance of the golden angle. In contrast, in the EDC2 parameter space, the region of intermediate openings is relatively narrowed and the region of openings near the golden angle is expanded, greatly increasing the dominance of the golden angle. These results support the validity of EDC2 and suggest that the protoplast age-dependent increase in inhibition is not limited to the case of cockscomb-type phyllotaxy, but is widely common to plants in general.
Our findings demonstrate the importance of temporal variation in interplastidial action in the regulation of phloem development and contribute to the elucidation of the nature of the phyllotaxis pattern. In the future, we expect that the accumulation of such studies will enable us to understand the various beautiful geometric patterns woven by nature from the viewpoint of generative principles.
Terminology:
Note 1: Protoplast
An organ in the early stages of formation that is immature and underdeveloped. The protoplast of a leaf is called the leaf primordium.
Note 2: Phyllotaxis
The arrangement of leaves surrounding a stem.
Note 3: Cockscomb-type phyllotaxy
A type of phyllotaxy with one leaf per node is called alternate. The kokusagi-shaped phyllotaxis is a form of alternate phyllotaxis, and is found in kokusagi-ginseng and other species. It is characterized by the regular and periodic changes in the direction of the leaves: 0°, 180°, 270°, 90°, 0°, and ……. In terms of opening, the change is from 180° to 90° to 180° to 270°. On branches that are not upright, there often appear to be two leaves on each side, such as left, left, right, right, ……, which is due to the twisting of the petiole.
Note 4: Divided tissue
A tissue that is responsible for the growth of plant organs through continuous cell division. The tissue at the tip of a stem is called shoot apical meristem or shoot apical meristem.
Note 5: Helical symmetry
A synthesis of rotation about an axis and translation along an axis.
Note 6: Two-row alternate phyllotaxy
In an alternate phyllotaxy, when the leaves are arranged in opposite positions at each node, this is called double-row alternate phyllotaxy. The degree of opening is 180°.
Note 7 Fibonacci spiral
A sequence of numbers such that the sum of two consecutive terms gives the next term, where the first two terms are 1, is called the Fibonacci sequence (1, 1, 2, 3, 5, 8, 13, …… and so on). An alternate phyllotaxy with openings close to the golden angle is called a Fibonacci spiral because it forms a beautiful helix, closely related to the Fibonacci sequence.
Note 8: Degree of opening
The angle between one leaf and the next leaf around the stem.
Note 9: Golden angle
The central angle of the shorter arc when a circle is bisected in such a way that the ratio of the total circumference to the length of the longer arc is equal to the ratio of the length of the longer arc to the length of the shorter arc, or the golden ratio. Approximately 137.5°.
Note 10: Crossed versus bifurcated
A crossed phyllotaxy is a phyllotaxy with two opposite leaves on a single node. Usually, in a paired phyllotaxy, the direction of leaf pairs changes by 90° at each node. This is called crossed opposite leaves.
Note 11: Three-lobed
A leaf stalk with three leaves per node. Usually, the three leaves are equiangular, i.e., 120° apart, and the three leaf pairs rotate 60° at each node.
Note 12: Parameter space
In the case of EDC2, there are five parameters, so the space is five-dimensional. In this study, we positioned the patterns of phyllotaxis output under each parameter condition in this space and examined the likelihood, stability, and mutual relationships of the various phyllotaxis based on their distribution.
Publication:
Journaj:「PLOS Computational Biology」(オンライン版:2019年6月6日)
Title:Mathematical model studies of the comprehensive generation of major and minor phyllotactic patterns in plants with a predominant focus on orixate phyllotaxis
Authors:Takaaki Yonekura, Akitoshi Iwamoto, Hironori Fujita, Munetaka Sugiyama